Materialien 3
5. Weiteres zur biologischen Evolution
a) über die Entstehung neuer Gene, b) über die Entstehung eines Bakterien-Motors.
a) W.Wieser (a.a.O., S.222) schreibt: Die zunehmende Differenzierung und Strukturierung
von Zellfunktionen (über die Einführung eines inneren Membransystems und Organellen
bei Eukarioten) ermöglichte größere Zellen und den Aufbau vielzelliger Organismen.
(Wenigzellige gibt´s merkwürdigerweise nicht, keine Zwei-, Drei- oder Zehnzeller!)
„Die Zunahme der Körpergröße und die konsequente Verwirklichung des Prinzips
der Differenzierung und Arbeitsteilung erlaubte die Evolution einer Mannigfaltigkeit
von Lebewesen, deren Repertoire an physiologischen Leistungen und Verhaltensweisen
um ein Vielfaches reicher war als das der bis dahin herrschenden Bewohner der
Biossphäre.“
Unsere Frage nun: Andersartige Wesen haben andere Gene. Wie ist die Entstehung
neuer Gene möglich?
Das DNS-Molekül hat die Fähigkeit, sich mit Hilfe von speziellen Polymerasen-Enzymen
so zu verdoppeln, dass die Nukleotidreihenfolge der Tochterstränge dieselbe
ist wie die der Matrix-Stränge. Es kann zu Kopierfehlern kommen: Mutationen.
Das ist der molekulare Mechanismus der Darwinschen Erbgutänderungen. Durch Mutation,
Selektion und Isolierung (Restabilisierung) sollen sich (der von Darwin begründeten
Evolutionstheorie zufolge) immer wieder neue Arten gebildet haben (Makroevolution)
und innerhalb der Arten Anpassungen, adaptive Veränderungen. Die Entstehung
neuer Arten setzt die Entstehung neuer Gene voraus. Neue Gene sind neue Kettenstücke
des Makromoleküls DNS. Ihre Entstehung, ihr Einbau, ist eine Polykondensation
genannte Synthesereaktion. Kann sie durch Mutation erfolgen? – Dazu wieder
Bruno Vollmert, die Fortsetzung seines bereits oben zitierten Radiovortrages
´Was Darwin nicht wissen konnte und Darwinisten nicht wissen wollen´. Seine
Antwort ist ´Nein´. Im folgenden die Begründung.
Da nach Ansicht der Darwinisten die DNS ein Lebewesen vollständig bestimmt,
kann man die Entwicklung eines Lebewesens von einem bestimmten Urbakterium zu
einem bestimmten Säugetier, wie lange es auch gedauert haben mag, mit der Synthese
seiner DNS im Laufe dieser Zeit gleichsetzen, einer statistischen (von selbst,
ohne äußere Eingriffe und Steuerungen wie im Labor oder in der Fabrik durch
den Chemiker verlaufenden) Polykondensation. Wir fragen nach der Wahrscheinlichkeit
für einer statistische, d.h. ´von-selbst´-Synthese.
Die Frage nach der historischen Entstehung neuer Lebewesenklassen ist damit
zur Frage nach der Entstehung eines Makromoleküls geworden.
Die Länge der DNS (verteilt auf Chromosomen) hat im Laufe der Zeit zugenommen:
beim Bakterium (und wie man annehmen kann wohl auch beim Urbakterium) beträgt
sie ca. 1mm, beim Säugetier ca. 1 m, Zunahme 1:1000 in drei bis vier Milliarden
Jahren. Die Menge der Gene stieg von wenigen Tausend auf 50 Tausend (das ist
die Zahl der im Säugetier vorkommenden verschiedenen Proteine, die ja von der
DNS codiert werden).
Neue Klassen von Lebewesen konnten nicht durch Mutation, also durch Veränderung
der Sequenz vorhandener DNS-Ketten entstehen, sondern nur durch Polykondensation,
d.h. durch Anwachsen zahlreicher neuer Gene an die DNS-Kette.
Mutationen ändern die Sequenz der Nukleotide in der DNS. (Sie Verlängern die
Kette nicht.) Nicht alle Mutationen machen
sich im Phänotyp (im Eigenschaftsbild) des Lebewesens bemerkbar, die allermeisten
bringen Defekte mit sich. Es kann aber auch eine günstige Eigenschaftsveränderung
sein, d.h. eine solche, die einen Fortpflanzungsvorteil (bei der Teilungsrate
oder der Nachkommenzahl bei sexueller Vermehrung) bilden. Dann greift die Selektion:
die eine Sorte stirbt allmählich aus, die andere lebt weiter.
Mutationen finden im Rahmen des vorhandenen Genoms statt. Neue Sequenzen sind
stets nur veränderte Vorhandene (ausgetauschte Vorhandene), keine zusätzlichen,
das Genom erweiternden.
Wie sich die DNS-Kette verlängerte, darauf kommt es nicht an, sondern nur darauf,
dass neue Information, neue Nukleotidsequenzen in Genlänge (ca. 1500 Nukleotide
in den vier Arten ATCG) entstanden bzw. eingebaut wurden.
Wie oben betont: Es geht um die Entstehung des DNS-Makromoleküls mit schließlich
50000 Genen (wie beim Säugetier) im Ausgang von einem DNS-Molekül mit nur ca.
1000 Genen (wie bei Bakterien). Statt mit dem historischen Nacheinander stets
komplexerer Lebewesen in der Evolution haben wir es jetzt mit dem gleichbedeutenden
sukzessiven Aufbau des DNS-Makromoleküls zu tun.
Die Reihenfolge des Wirksamwerdens der ca. 50000 Gene des Säugetiers ist zu
bedenken: Ein Gen bzw. das ihm entsprechende Enzym, dass beim Säugetier für
das Wachstum des Fingernagels nützlich ist, ist im Entwicklungsstadium (auf
der Evolutionsstufe) des Wurmes wenig hilfreich. Dessen Gene müssen vor denen
des Säugetiers entstehen. Allgemein gilt: Ein neues Gen muss mit den bereits
vorhandenen kooperieren, um die Funktionsfähigkeit des Organismus zu gewährleisten.
Von den 50 Tausend Säugetier-Genen war in einer gegebenen Evolutionssituation
immer nur eine Sequenz brauchbar. Die Wahrscheinlichkeit der Entstehung ist
dann 1:50000. Allerdings gibt es ca. 10 hoch 700 mögliche Nukleotidsequenzen
in Genlänge (also 10 hoch 700 mögliche Gene statt der nur 50000 tatsächlichen).
Wir lassen das aber jetzt außer Betracht. Es reicht die (Un)Wahrscheinlichkeit
von 1:50000.
Durchweg bewirkt ein einziges neues Gen in einer Zelle nichts. Für die Synthese
eines physiologisch wirksamen Stoffes sind meist ca. 10 Enzyme nötig, d.h. ca.
10 Gene. Eigenschaftsveränderungen bedürfen meist der Zuschaltung von 30 bis
50 Enzymen bzw. Genen. (Mutationen, die vorhandene Gene verändern, wirken meist
drastisch schon bei der Veränderung nur eines Gens.) Eine Selektion kann also
bei der Polykondensation (beim Anfügen noch nicht im Betrieb befindlicher, erst
im Verbund mit vielen anderen neuen Genen wirksamer Gene)
nicht greifen, weil ein neues, zusätzliches Gen sich durchweg nicht im Eigenschaftsbild
(d.h. hier: als Fortpflanzungsvorteil oder Nachteil) bemerkbar macht. Das Leben
geht (anders als bei der Mutation eines vorhandenen Genes) ungestört weiter.
Es gibt keine Testmöglichkeit für das neue Gen. Nur neue Phänotyp-Eigenschaften
sind testbar.
In der Industrie macht man von Optimierungsprozessen durch zufällige Systemvariationen
Gebrauch (sog. Evolutions-Experimente). Das entspricht dem Testen von Mutationen.
In der Natur geschieht das auch. Aber nur im Sinne der Optimierung (besserer
Anpassung) einer bereits vorhandenen Konstruktion. Das führt nicht zu Entstehung
neuer Arten (zu neuen Gen-Komplexen in der DNS). Baut man ein Auto in ein Flugzeug
um, kann man die einzelnen Konstruktions- und Montageschritte ebenso wenig testen,
wie es bei der Addition neuer Gene in der Natur möglich ist.
Die Addition eines passenden Gens hat die Wahrscheinlichkeit 1:50000, die von
ca. 20 Genen, die zusammen z.B. die Glykocholsäure synthetisieren, 1:10 hoch
100. Geht man von eine 5-stufigen Evolution aus (1. Ursuppe bis Zelle, 2. Zelle
bis Wirbellose, 3. Wirbellose bis Fische, 4. Fische bis Reptilien, 5. Reptilien
bis Säuger), so bedeutet jede Stufe einen Längenzuwachs der DNS von ca. 10000
Genen. Die Wahrscheinlichkeit des Übergangs durch zufälliges Kettenwachstum
ist dann gleich ca. 1: 10 hoch 40000. Hätte jede Stufe 20 Mittelglieder, bestünde
für jeden der Schritte noch die (Un)Wahrscheinlichkeit 1 durch 10 hoch 2000.
Man schätzt die Zahl der Atome im gesamten Universum auf 10 hoch 80. Wenn dann
das ganze Universum aus Nukleinsäuremolekülen bestände, wäre die Chance, darin
auch nur eine zufällig entstandene DNS-Kette mit 4 oder 5 Hundert mit einander
(im Sinne der Bildung von Enzymketten) kooperierenden) Genen zu finden (d.h.
dass eine solche dabei wäre), immer noch unvorstellbar gering. Oder anders:
Es müsste 10 hoch 2000 Reptilien gegeben haben (mehr als Atome im Universum),
damit unter ihnen eines das erste von 20 lebensfähigen Urvogel-Zwischenstufen
hätte sein können.
Bruno Vollmert: Evolution als statistische (ungesteuerte) Polykondensation
kann es nicht gegeben haben. Alternative: gesteuerte Polykondensation! –
Durch den göttlichen Chemiker? Also Schöpfung statt Evolution.
b) Darwin schreibt 1859 in Über die Entstehung der Arten: „Wenn
bewiesen werden könnte, dass irgendein komplexes Organ existiert, das unmöglich
durch viele aufeinanderfolgende kleine Veränderungen entstanden sein kann, würde
meine Theorie vollkommen zusammenbrechen.“ Sie ist zusammengebrochen.
Der Beweis wird im folgenden erbracht für das Organ ´Antriebsmotor´ des Bakterium
Escherichia coli.
Ein irreduzibel komplexes Organ ist ein System, dass aus mehreren in Wechselbeziehung
zueinander stehenden Teilen besteht, wobei die Entfernung auch nur eines Teiles
die Funktion des ganzen Systems zerstört – wie bei einer Mausefalle. Solche
System können sich unmöglich durch den Darwinschen Prozess (Mutation, Selektion)
entwickeln, weil nur das ganze System funktionsfähig ist und alle Vorstufen
durch Selektion als funktionsunfähig eliminiert werden. (Der Gegensatz zur irreduziblen
Komplexität ist kummulative Komplexität.) In der Natur gibt es viele molekulare
Maschinen, die irreduzible Komplexität besitzen, zum Beispiel der kleine Elektromotor,
durch den sich Bakterien im Wasser fortbewegen können. 35000 solcher Motoren
sind gerade mal einen Millimeter lang! – Die Wahrscheinlichkeit für die
Entstehung eines Motors, und sei es auch eine angenommene primitivere Form als
die bestehende, ist, wie im folgenden belegt wird 1:10 hoch 29. Ich beziehe
mich auf Siegfried Scherer / Laurence Loewe, ´Probleme der Erklärung molekularer
Maschinen durch Evolution´ in: Paul Weingartner (Hg.), Evolution als Schöpfung.
Kohlhammer: Stuttgart Berlin Köln 2001.
Zahlreiche Bakterien können sich aktiv bewegen (Richtung Nährstoffquellen,
Ausweichen vor Gefahrenquellen). Manche verfügen über Rotationsmotoren zur Geißelbewegung.
(Abb.1)
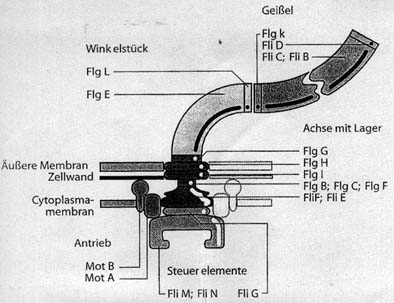
Abb. 1: Schemazeichnung einer Bakterienzelle mit Rotationsmotor
und Geißel. Das Feld am "Vorderende" des Bakteriums bezeichnet
einen Bereich der Cytoplasmamembran, der dicht mit Chemosensoren besetzt ist.
Man hat dieses Chemosensorenfeld auch die "Nase" des
Bakteriums genannt. Von dort werden Steuersignale (Pfeile) an die Motoren übertragen,
die ihrerseits die Flagellen in Rotation versetzen.
Flagellen erzeugen durch die Rotation den Vortrieb. Nach (Maddock et al. 1993)
und (Parkinson et al. 1993), verändert.
Der Motor bei Escherichia coli wird von 40 Proteinen gebildet. Die Steuerung
erfolgt durch ca. 10 Proteine. Die zuständigen Gene enthalten 60000 Basenpaare.
Ein Motor besteht aus fünf Funktionselementen: Geißel, Winkelstück, Rotationsachse,
Lager, Motorprotein. (Abb.2)
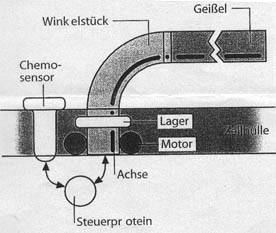
Abb. 2: Schemazeichnung der Hauptstrukturkomponenten
eines Bakterienmotors. Die Bestandteile der Geißel, der Winkelstücks,
der Achse mit den den Lagern, sowie des Antriebskomplexes sind mit den Abkürzungen
für die Namen der entsprechenden Gene versehen.
Der Motor ist mit Gramnegativen Bakterien zwischen äußerer Membran, Zellwand
und Cytoplasmamembran lokalisiert. (Nach (Falke et al. 1995), verändert.
Die Geißel (gewissermaßen die Schiffsschraube), aufgebaut durch das Protein
Flagellin, bestehend aus über 400 Aminosäuren, ist über ein Winkelstück mit
der Rotationsachse verbunden, deren Lager von der Cytoplasma-Membran und Zellwand
gehalten wird. Die Gene für (die Proteine für) Achse und Lager sind bekannt.
Auch die Gensequenz für das Rotieren. Aber wie letzteres erfolgt, weiß man nicht.
Der Motor wird elektrisch betrieben, durch die 0,2 Volt Membranpotential, die
das Gesamtbakterium (die Zelle) als Batterie erzeugt. Der Motor verbraucht ca.
1200 Protonen pro Umdrehung mit der Höchstgeschwindigkeit von 50 mm pro sec
(entspricht 450 km/h beim Auto).
Angenommen, es gab einen Evolutionsprozess. Wie konnte ein solches Organ, der
Bakterienmotor (eine dieser Einzeller hat 5 bis 10 solcher Motoren), entstehen?
Gab es Zwischenstufen, funktionierende einfacherer Motoren? Wie könnte der einfachste
Motor ausgesehen haben? Wie groß war die Wahrscheinlichkeit für das Entstehen
des angenommen einfachsten Motors? (Abb.3)
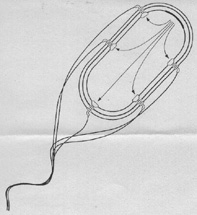
Abb. 3: Notwendige Komponenten für einen hypothetischen,
"primitiven" Bakterienrotationsmotor.
Jede Komponente ist notwendig, damit eine erste selektionsfähige Struktur entsteht.
Einzelheiten im Text.
Ein einfacherer und bzgl. des Zustandekommens wahrscheinlicherer Motor müsste
auch die 5 Grundelemente gehabt haben. Würde eines fehlen, wäre er nicht funktionsfähig
gewesen. Die entstandene Struktur ohne dies Element wäre lediglich eine Belastung
für das Bakterium gewesen, es hätte im Selektionsprozess nicht bestehen können
und wäre ausgestorben. Nehmen wir an, 5 Proteine reichten für die fünf Elemente
(beim tatsächlichen Bakterienmotor sind es 40). Für die Steuerung nehmen wir
dann noch zwei Proteine (statt der tatsächlichen 10). Wir nehmen dann auch noch
an, dass für die neuen Proteine umzubauende Vorläuferproteine bestanden haben
und dass sie so ähnlich waren, dass nur drei Aminosäurepositionen neu besetzt
werden mussten. Weiter: dass der Umbau in Duplikaten von Genen (sagen wir nur
sieben bei dem angenommenen Einfachmotor) stattgefunden hat (so dass der bestehende
Organismus funktionsfähig blieb, das er ja nicht „wegen Umbau vorübergehend
geschlossen“ werden konnte). Auch sollten die präadaptierten Genduplikate
während ihres Umbaus nicht abgeschaltet sein. Die Motormutationen sollten also
nicht in abgeschalteten Genen stattgefunden haben.
Für einen neuen Funktionszustand mit den wenigsten Veränderungen braucht man
dann sieben veränderte Proteine. - Nun können nicht wenige Aminosäuren in einem
Protein verändert werden, ohne nennenswerte Funktionsänderung hervorzurufen.
Und es gibt Mutationen, die sehr starke Änderungen (manchmal sogar Verbesserungen
– letztlich hinsichtlich der Vermehrungsrate) hervorrufen. Nehmen wir
an, es reichten drei Veränderungen, um einem Protein von 100 Aminosäuren eine
neue Funktion zu geben.
Die Wahrscheinlichkeit für eine Mutation bei einem Gen von 1000 Basenpaaren
beträgt 1:10 hoch 5. D.h.: unter 100 000 Bakterien trägt eines seiner Gene an
einer beliebigen Stelle (tatsächlich müsste es an einer bestimmten sein) eine
Mutation mit der Wahrscheinlichkeit von ca. 0,6. Soll in einer Generation ein
Gen in drei Schritten (Mutationen) zu einem Gen für ein Bakterienmotorprotein
umgebaut werden, so ist dafür die Wahrscheinlichkeit 1:10 hoch 15. Für einen
spürbaren Selektionsvorteil (durch funktionierenden 5-teiligen Motor) brauchen
wir je drei Mutationen an sieben Genen für sieben Proteine (5 Motorproteine,
2 Steuerproteine), macht 21 Mutationen – an 7 Genduplikaten, für die 7
Veränderungen (Duplikationen) nötig sind, macht 28 unabhängigeVeränderungen
insgesamt. Ihre Wahrscheinlichkeit beträgt 1:10 hoch 140 pro Generation pro
Zelle.
Zu einem gegebenen Zeitpunkt gibt es auf der Erde höchstens 10 hoch 32 Zellen.
Die Wahrscheinlichkeit für die Entstehung eines Motors in einer Generation auf
der ganzen Erde liegt dann bei 1:10 hoch 108.
Ein Beweis für die Unmöglichkeit der Makroevolution ist das noch nicht, wohl
dafür, dass sie mit den bisherigen Prinzipien Mutation und Selektion nicht vonstatten
gehen konnte. Makroevolution, die Entstehung neuartiger molekularer Maschinen
oder neuartige Baupläne für Organismen (neue Arten!), konnte bisher experimentell
nicht beobachtet werden. Für das Problem ihrer Entstehung gibt es keine mathematisch
exakte begründete Antworten (Scherer / Loewe a.a.O. S. 181). Mikroevolutive
Vorgänge sind meist Optimierungsvorgänge.
Die Darwinsche Evolution erfolgt über Mutation mit anschließender Selektion
der Genotypen, welche die höchste Fortpflanzungsrate unter den gegebenen Umweltbedingungen
aufweisen. Die gibt es natürlich. Aber dabei entstehen keine neuartigen Konstruktionen
– wie z.B. ein Bakterienmotor.
Zurück
zur Auswahl